Lin Chao
Research
We use a combination of computer modeling and laboratory experimentation to study ecological and evolutionary processes. Ecological and evolutionary studies are often difficult to investigate because they occur slowly over long periods of time. Computer models can speed up time, but we also take advantage of the fact that microbes, bacteria and viruses provide that same advantage because of their short generation times. Furthermore, they can be easily cultured in the laboratory under controlled conditions. However, we also realize that too much control can be dangerous because an experiment that is too controlled becomes artificial and not much more than a simulation. And a simulation is always more easily obtained from a computer model. Thus, we are careful to design experiments that teach us more than a computer model. We make sure that the experiments allow us to measure an unknown that could not be determined a priori from a computer model. We have used microbial systems to study topics ranging from the ecology of host-parasite co-existence and the evolution of sex, mutation rates, transposable elements, group and individual adaptations, game theory strategies, Fisher’s Geometric Model of Adaptive Evolution, deleterious and compensatory mutations. As new information or interests develop, we often revisit older topics or explore new ones.
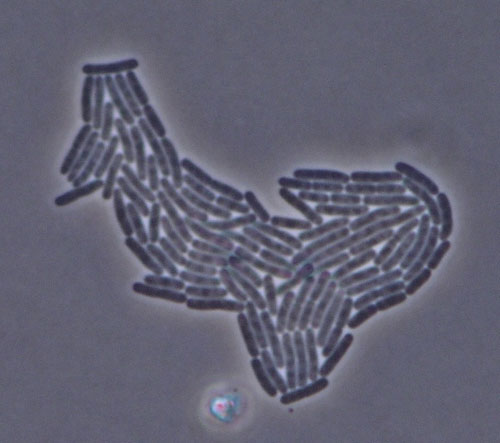
Time lapse photograph E. coli cells descending from a single cell. By recording division of all cells via time lapse, the doubling time or life history fitness of every individual in a population can be determined.
Our current research focuses on evolution of microbial aging or senescence, which grew out of our interest in genetic damage or mutations. Because most mutations are deleterious, they lead to fitness loss unless removed by natural selection. The transmission of mutations from mother to daughter is governed by well the well known rules of genetic inheritance and assortment. However, non-genetic or phenotypic damage can also decrease fitness and, unlike mutations, the transmission rules for phenotypic damage are not well understood. If a mother cell has ten units of phenotypic damage, for example ten proteins damaged by oxidation, should she distribute the damage evenly between her two daughters or give all ten units to one daughter and none to the other? Our theoretical models have shown that a lineage maximizes its fitness by allocating all the damage to a single daughter. Thus, asymmetry should evolve and the daughter receiving all or more damage should senesce over time. Time lapse photography of dividing cells has already shown that bacteria divide asymmetrically. We are now making our own time lapse series to test more directly the prediction of our models.
Select Publications
- Liao, M. J., A. Miano, C. B. Nguyen, L. Chao, J. Hasty. 2020. Survival of the weakest in non-transitive asymmetric interactions among strains of E. coli. Nat Commun 11:6055.
- Shi C, Chao L, Proenca AM, Qiu A, Chao J, Rang CU. 2020. Allocation of gene products to daughter cells is determined by the age of the mother in single Escherichia coli cells. Proceedings Royal Society B 287:20200569.
- Proenca AM, Rang CU, Qiu A, Shi C, Chao L. 2019. Cell aging preserves cellular immortality in the presence of lethal levels of damage. PLoS Biol 17(5): e3000266. https://doi.org/10.1371/ journal.pbio.3000266
- A. M. Proenca, C. U. Rang, C. Buetz, C. Shi & L. Chao. 2018. Age structure landscapes emerge from the equilibrium between aging and rejuvenation in bacterial populations. NATURE COMMUNICATIONS 9:3722. DOI: 10.1038/s41467-018-06154-9
- Rang CU, Proenca A, Buetz C, Shi C, Chao L. 2018. Minicells as a damage disposal mechanism in Escherichia coli. mSphere 3:e00428-18. https://doi.org/10.1128/mSphere.00428-18.
- Chao L, Elena SF. 2017. Nonlinear trade-offs allow the cooperation game to evolve from Prisoner’s Dilemma to Snowdrift. Proc. R. Soc. B 284: 20170228. http://dx.doi.org/10.1098/rspb.2017.0228
- Chao, L., Rang, C. U., Proenca, A. M. and Chao, J.U. 2016. Asymmetrical Damage Partitioning in Bacteria: A Model for the Evolution of Stochasticity, Determinism, and Genetic Assimilation. PLOS Computational Biology DOI: 10.1371/journal.pcbi.1004700
- Campbell EM, Chao L. 2014. A Population Model Evaluating the Consequences of the Evolution of Double-Resistance and Tradeoffs on the Benefits of Two-Drug Antibiotic Treatments. PLoS ONE doi:10.1371/journal.pone.0086971
- Samuel L Díaz-Muñoz, Olivier Tenaillon, Daniel Goldhill, Kristen Brao, Paul E Turner and Lin Chao. 2013. Electrophoretic mobility confirms reassortment bias among geographic isolates of segmented RNA phages. BMC Evolutionary Biology 13:206.
- Rang, C. U., Peng, A. Y. and Chao, L. 2012. Bacterial aging requires an extrinsic damage agent. Microbiology 158:1553–1559
- Rang, C. U., A. Y. Peng, and L Chao. 2011. Temporal Dynamics of Bacterial Aging and Rejuvenation. Current Biology. In Press.
- Chao L. 2010. A Model for Damage Load and Its Implications for the Evolution of Bacterial Aging. PLoS Genet 6(8): e1001076. doi:10.1371/journal.pgen.1001076
- O’Keefe, K. J., O. K. Silander, H. McCreery, D. M. Weinreich, K. M. Wright, L. Chao, S. V. Edwards, S. K. Remold, and P. E. Turner. 2010. Biogeography of sexual reassortment in RNA phage. Evolution. doi:10.1111/j.1558-5646.2010.01040.x.
- Tenaillon, O., Silander, O., Uzan, J-P., and L. Chao. 2007. Quantifying Organismal Complexity using a Population Genetic Approach. PLoS ONE 2:1-8.
- Olin K. Silander, Olivier Tenaillon and Lin Chao. 2007. Understanding the Evolutionary Fate of Finite Populations:The Dynamics of Mutational Effects. PLoS Biol 5:922-931.
- Ackermann, M., Chao, L, Bergstrom, C. T., and Doebeli, M. 2007. On the Evolutionary Origin of Aging. Aging Cell 6:235-244.
- Poon, A. and L. Chao. 2006. Functional origins of fitness-effect sizes of compensatory mutations in the DNA bacteriophage PhiX174. Evolution 60: 2032-2043.
- Chao, L., C. U. Rang, and L. E. Wong. 2002. The distribution of spontaneous mutants and inferences about the replication mode of the RNA bacteriophage φ6. J. of Virology 76:3276–3281.
- Burch, L. C. and L. Chao. 2000. Evolvability of an RNA virus determined by its mutational neighborhood. Nature 406:625-628.
- Turner, P. E. and L. Chao. 1999. Prisoner’s Dilemma in an RNA Virus. Nature 398:441-443.
Biography
Lin Chao received his Ph.D. from the University of Massachusetts, Amherst. He was an NIH postdoctoral fellow at Princeton Universit/y./ Lin joined UCSD in 1999 and he served as the Chair of the Section of Ecology, Behavior and Evolution from 2004 to 2008. In 2014, he received the UCSD Chancellor's Associates Award for Excellence in undergraduate teaching.
